Problem set #3. Analog/digital axonal transmission.
In one of this week's papers, Shu et al. (2006) showed that a significant fraction of cell body membrane potential changes, for example induced by synaptic input, were experienced by the axon, and they calculated an axonal membrane space constant (l) of ~ 500 mm. They proposed that the delivery of subthreshold (non-spike) signals to the axon and its synaptic terminals resulted in a modification of synaptic efficacy Ц i.e. that excitatory postsynaptic synaptic potentials were increased in amplitude when the neuron sending the signal (the presynaptic neuron) was depolarized in the steady state. This provocative result indicates that encoding of information in neural networks has second order features Ц i.e. that the net signal received by a neuron depends not only the mean spike frequency of the presynaptic neuron, but also on the general state of excitability of the neuron.
A key concept discussed in class is that of membrane space constant, which can be calculated as:
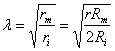
In NEURON,
r (radius) is: diam (um)/2
Rm is: 1/g_pas(S/cm2)
Ri is: Ra (ohm-cm)
1) Obtain the zipped archive file for the NEURON code for the Shu paper here ps3.zip. Load the best_full_axon_decay.hoc file into neuron. What are the dimensions, space constant, and time constant of the initial segment? Graph[1] of the simulation plots the voltage along several sections of the neuron, including an axon branch. While there are many axon branches generated in the .hoc file, only one is plotted in Graph[1]. You can find the plotting code after the second instance of тАЬnew Graph(0)тАЭ in the .hoc file. Write out the name and length of each section being plotted тАУ be sure it adds up to what is being plotted on the graph. You may want to look up the RangeVarPlot function on the Neuron ProgrammerтАЩs Reference, and also see how the first few sections are connected in the procedure тАЬcreate_axon_connection()тАЭ. Calculate the space constant of the last section (the axon) being plotted.
2) Run the simulation, which provides some noisy synaptic input into the soma and dendrites of the neuron, and measures voltages in the soma, initial segment, and throughout the axon as described in 1. An output file (seg.dat) is created for sampled locations along the axon: every 5 u m for the first 50 um and 7.5 um after that. The first column in the table is sample time points, and the subsequent columns are the recorded voltages in each of the locations. Using this output file calculate the empirical space constant as the point at which the somatic (x = 0, at the beginning of the axon) response (i.e. the change in voltage from rest produced by the noisy synaptic input) decays by 1/e. Is this empirical space constant the same as either of the theoretical constants from problem #1? Explain. At what noise level do you start to see spiking?
3) Let's mimic the specific effect of a certain class of inhibitory neurons, so-called chandelier cells, which specifically target the axon initial segment, upon axonal function. At equilibrium, we can mimic this by increasing the leak conductance to reproduce the effect of the chandelier cell output. Assume for now that the channels opened by the chandelier cells have the same reversal potential as the resting conductance (g_pas). If a chandelier cell makes 30 contacts onto an axon initial segment, and each contact mediates an increase of 0.4 nS, how would you modify the properties of the iseg to mimic the continuous activation of chandelier cell output? What is the effect of this manipulation on the space and time constant of the initial segment? What about the effect on the empirical space constant in the total axon, as determined in step 2? At what noise level do you start to see spiking with this manipulation? Explain.